Sex difference in the profile of extracellular bioactive lipids of conjunctival epithelial cells during allergic inflammation
The ocular surface is comprised of cornea and conjunctiva overlaid by the tear film1. The epithelium of the conjunctiva lines the inside of the eyelids and covers the sclera, thus providing stability to the transparent cornea and anterior segment of the eye. The goblet cells in the conjunctival epithelium (CjGCs) secrete water, electrolytes, and mucins to wet and protect the ocular surface. Under- or over-secretion of mucins due to inflammation causes ocular surface instability, visual disturbance, and disease2, 3.
Allergic conjunctivitis (AC) is an allergic inflammation developed in conjunctival epithelium, which is one of the most common ocular allergies affecting more than 40 % of the North American population4. Symptoms caused by AC include pruritis (itchy eyelid skin), excess tearing, conjunctival redness, foreign body sensation, mucous discharge, and eyelid swelling. Many individuals with AC also experience allergic rhinitis, asthma, eczema, food allergy, and eosinophilic esophagitis, which make daily life even more debilitating. The molecule that causes multiple types of allergic reactions is histamine. Released immediately after allergen sensitization by mast cells, histamine binds to histamine receptors (H1, H2, H3, and H4) on the mast cell surface to promote the production and secretion of proinflammatory mediators prostaglandins, leukotrienes, and thromboxanes, which collectively enhance the inflammatory response. Histamine binding to histamine receptors in CjGCs increases the intracellular Ca2+ concentration ([Ca2+]i), activates ERK1/2, and induces mucin over-secretion in CjGCs5.
At the peak of the inflammatory response, various specialized pro-resolving mediators (SPMs) start to sequentially appear at the site of inflammation. SPMs are a class of bioactive lipid mediators derived from omega-3 (ω-3) or ω-6 polyunsaturated fatty acids (PUFAs) that resolve inflammation. These SPMs include ω-3 PUFA-derived resolvins, maresins, protectins, and ω-6 PUFA-derived lipoxins6. SPMs provided by infiltrating immune cells (i.e., neutrophils, monocytes, and macrophages) collectively resolve inflammation by inhibiting apoptotic polymorphonuclear infiltration, neutrophil trans-epithelial migration, and proinflammatory cytokine release by epithelial cells, as well as by promoting macrophage efferocytosis. These extracellular SPMs also counter-regulate histamine-mediated mucin oversecretion in CjGCs7-10.
The onset of an allergic reaction and its resolution are relatively well studied from the infiltrating immune cell standpoint; thus, the current regimen for treating allergic conjunctivitis (i.e., ocular antihistamine agents, mast cell stabilizers, ophthalmic steroids, and non-steroidal anti-inflammatory drugs) is focused on regulating immune cell activity or mitigating the action of inflammatory mediators4. Despite advancements in our understanding and treatment options for allergic conjunctivitis, how conjunctival epithelial cells respond to allergic inflammation is still unknown. More specifically, there is no report on whether conjunctival epithelial cells only indirectly respond to the extracellular mediators provided by infiltrating immune cells, thus merely controlling the level of mucin secretion from CjGCs or take active measures to resolve inflammation, such as by biosynthesizing SPMs.
We targeted our investigation to lipid mediators in extracellular vesicles (EVs). EVs are cell-secreted nano-vesicles made up of a lipid bilayer and carry nucleic acids, proteins, and lipids11-13. Due to their ability to mediate cell-to-cell communication in an auto-, para-, and endocrine manner, EVs carry different molecules that reflect surrounding biological condition. This dynamism makes them an important biological entity to investigate and understand health and disease better.
We report here that human primary CjGCs (HCjGCs) biosynthesize lipid mediators (proinflammatory mediators and SPMs), load them into EVs, and secrete to the extracellular space. At 18 hr post histamine stimulation, the amount of certain lipid mediators significantly increased only in female EVs, but not in male EVs (Figure 1).
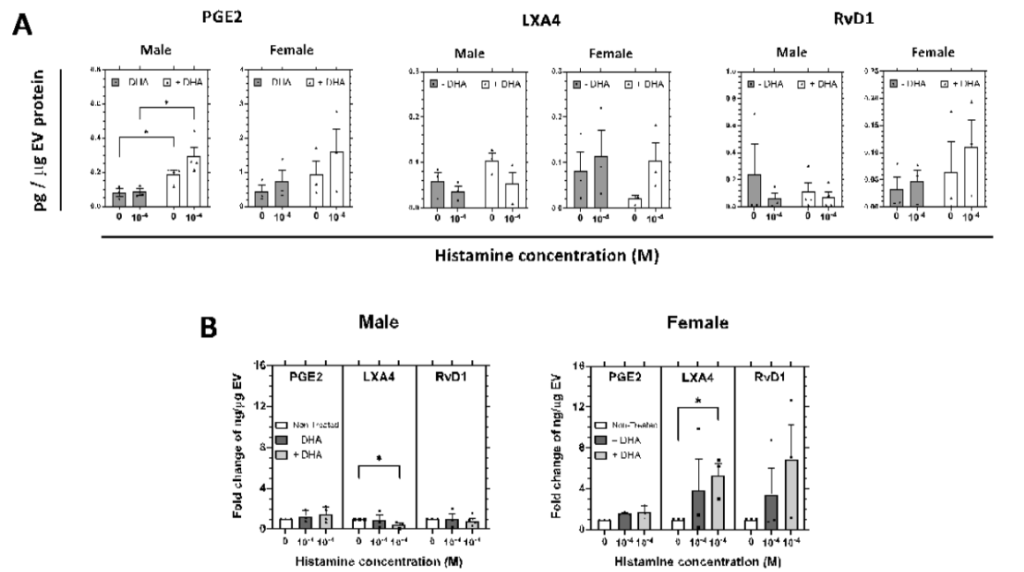
In response to histamine, the amount of RvD1, LXA4, and PGE2 increased in female EVs (EVs secreted from female HCjGCs) in general, but no trend was observed in male EVs (EVs secreted from male HCjGCs) (Figure 1A, grey bars 0 vs. 10-4). We then tested if supplementing docosahexaenoic acid (DHA) can increase the amount of SPMs in male EVs. DHA is a type of ω-3 PUFA, a precursor to three families of SPMs: resolvins (includes RvD1), protectins, and maresins14. In female EVs, the addition of DHA resulted in a higher amount of RvD1 at both basal and histamine stimulation, whereas males show no difference in RvD1 (Figure 1A, white bars vs. grey bars in RvD1). The addition of DHA even led to decreased LXA4, which is not a derivative of DHA, amount in female EVs under basal conditions, likely at the expense of increased RvD1 amount, but its amount was appropriately increased upon histamine stimulation (Figure 1A, white bars vs. grey bars in LXA4). The addition of DHA elevated LXA4 in male EVs under basal conditions, but it did not increase further by histamine stimulation. The addition of DHA affected the amount of PGE2 in both male and female EVs. In male EVs, there was no difference in the amount of PGE2 between basal and histamine stimulation without DHA, but the amount of PGE2 was increased in general and showed a difference between basal and histamine stimulation in the presence of DHA. This trend was also observed in female EVs (Figure 1A, white bars vs. grey bars in PGE2).
Since the HCjGCs were expanded ex vivo from deceased donors with very different medical conditions, it would be more intuitive to visualize the quantification in fold change. Without DHA, RvD1 and LXA4 amounts were 3~4 fold higher in female EVs, but no change was observed in male EVs (Figure 1B, dark grey bar vs. white bar). In the presence of DHA, female EVs showed even higher RvD1 (6~7 fold) and LXA4 (5~6 fold), suggesting enhanced pro-resolving capability in females (Figure 1B, light grey bar vs dark grey bar). This fold increase was observed only in female EVs but not in males. There was almost no fold change in male EVs except a significant decrease in LXA4.
Our result expands our understanding of CjGC physiology beyond mucin secretion. It proved the existence of active measures that these cells take during allergic nflammation and also shows a notable sex difference in allergic response between males and females. We also hope that our result gives insights into explaining the male predisposition (male-to-female ratio 3:1) to severe forms of conjunctivitis, i.e., vernal keratoconjunctivitis and atopic keratoconjunctivitis, observed in the clinic and the better treatment.
References
1. Craig JP, Nichols KK, Akpek EK, Caffery B, Dua HS, Joo CK, Liu Z, Nelson JD, Nichols JJ, Tsubota K, Stapleton F. TFOS DEWS II Definition and Classification Report. Ocul Surf. 2017;15(3):276-83. Epub 2017/07/25. doi: 10.1016/j.jtos.2017.05.008. PubMed PMID: 28736335.
2. Dartt DA, Masli S. Conjunctival epithelial and goblet cell function in chronic inflammation and ocular allergic inflammation. Curr Opin Allergy Clin Immunol. 2014;14(5):464-70. Epub 2014/07/26. doi: 10.1097/ACI.0000000000000098. PubMed PMID: 25061855; PMCID: PMC4206188.
3. Hodges RR, Dartt DA. Tear film mucins: front line defenders of the ocular surface; comparison with airway and gastrointestinal tract mucins. Exp Eye Res. 2013;117:62-78. Epub 2013/08/21. doi: 10.1016/j.exer.2013.07.027. PubMed PMID: 23954166; PMCID: PMC4222248.
4. Dupuis P, Prokopich CL, Hynes A, Kim H. A contemporary look at allergic conjunctivitis. Allergy Asthma Clin Immunol. 2020;16:5. Epub 2020/01/30. doi: 10.1186/s13223-020-0403-9. PubMed PMID: 31993069; PMCID: PMC6975089.
5. Hayashi D, Li D, Hayashi C, Shatos M, Hodges RR, Dartt DA. Role of histamine and its receptor subtypes in stimulation of conjunctival goblet cell secretion. Invest Ophthalmol Vis Sci. 2012;53(6):2993-3003. Epub 2012/04/03. doi: 10.1167/iovs.11-8748. PubMed PMID: 22467574; PMCID: PMC3376076 R.R. Hodges, None; D.A. Dartt, None.
6. Chiang N, Serhan CN. Structural elucidation and physiologic functions of specialized pro-resolving mediators and their receptors. Mol Aspects Med. 2017;58:114-29. Epub 2017/03/25. doi: 10.1016/j.mam.2017.03.005. PubMed PMID: 28336292; PMCID: PMC5623601.
7. Dartt DA, Hodges RR, Li D, Shatos MA, Lashkari K, Serhan CN. Conjunctival goblet cell secretion stimulated by leukotrienes is reduced by resolvins D1 and E1 to promote resolution of inflammation. J Immunol. 2011;186(7):4455-66. Epub 2011/03/02. doi: 10.4049/jimmunol.1000833. PubMed PMID: 21357260; PMCID: PMC3532815.
8. Hodges RR, Li D, Shatos MA, Serhan CN, Dartt DA. Lipoxin A(4) Counter-regulates Histamine-stimulated Glycoconjugate Secretion in Conjunctival Goblet Cells. Sci Rep. 2016;6:36124. Epub 2016/11/09. doi: 10.1038/srep36124. PubMed PMID: 27824117; PMCID: PMC5099697.
9. Li D, Hodges RR, Jiao J, Carozza RB, Shatos MA, Chiang N, Serhan CN, Dartt DA. Resolvin D1 and aspirin-triggered resolvin D1 regulate histamine-stimulated conjunctival goblet cell secretion. Mucosal Immunol. 2013;6(6):1119-30. Epub 2013/03/07. doi: 10.1038/mi.2013.7. PubMed PMID: 23462912; PMCID: PMC3742576.
10. Serhan CN. Discovery of specialized pro-resolving mediators marks the dawn of resolution physiology and pharmacology. Mol Aspects Med. 2017;58:1-11. Epub 2017/03/07. doi: 10.1016/j.mam.2017.03.001. PubMed PMID: 28263773; PMCID: PMC5582020.
11. Kalluri R, LeBleu VS. The biology, function, and biomedical applications of exosomes. Science. 2020;367(6478). Epub 2020/02/08. doi: 10.1126/science.aau6977. PubMed PMID: 32029601; PMCID: PMC7717626.
12. Mathieu M, Martin-Jaular L, Lavieu G, Thery C. Specificities of secretion and uptake of exosomes and other extracellular vesicles for cell-to-cell communication. Nat Cell Biol. 2019;21(1):9-17. Epub 2019/01/04. doi: 10.1038/s41556-018-0250-9. PubMed PMID: 30602770.
13. van Niel G, D’Angelo G, Raposo G. Shedding light on the cell biology of extracellular vesicles. Nat Rev Mol Cell Biol. 2018;19(4):213-28. Epub 2018/01/18. doi: 10.1038/nrm.2017.125. PubMed PMID: 29339798.
14. Spite M, Claria J, Serhan CN. Resolvins, specialized proresolving lipid mediators, and their potential roles in metabolic diseases. Cell Metab. 2014;19(1):21-36. Epub 2013/11/19. doi: 10.1016/j.cmet.2013.10.006. PubMed PMID: 24239568; PMCID: PMC3947989.
Reported by Dr. Changrim Lee (2023 KSEA YIG Grant Recipient)

